
| RNA-polymerase II | ||||
|---|---|---|---|---|

| ||||
|
RNA-polymerase II van Saccharomyces cerevisiae
| ||||
| Identificatie | ||||
| Alt. naam | DNA-Directed RNA Polymerase II | |||
| EC-nummer | 2.7.7.6 | |||
| CAS-nummer | 9014-24-8 | |||
| Databanken | ||||
| IntEnz | IntEnz view | |||
| BRENDA | BRENDA entry | |||
| ExPASy | NiceZyme view | |||
| KEGG | KEGG entry | |||
| MetaCyc | Stofwisselingsroute | |||
| PDB-codes | ||||
| ||||

De RNA-polymerase II, RNAP II, Pol II of RNA-polymerase B is een RNA-polymerase, die betrokken is bij de transcriptie van de mRNA's, de meeste snRNA's en microRNA's. Het enzym komt voor in het kernplasma van eukaryoten en bestaat uit 10 tot 12 subeenheden. De RNA-polymerase II van de mens heeft 12 subeenheden. RNA-polymerase II bestaat uit de vier mobiele elementen: Core, Clamp, Shelf en Jaw Lobe, die ten opzichte van elkaar kunnen bewegen. Het Core-element bestaat uit RPB3, 10, 11, 12 en delen van RPB1 en RPB2, die het actieve centrum vormen. Dit element heeft ongeveer de helft van de massa van RNA-polymerase II.
De wetenschappers Robert Roeder en William Rutter ontdekten in 1969 een derde RNA-polymerase. Ze gebruikten DEAE-Sephadex (diethylaminoethylcellulose; een ionenwisselaarhars) bij de ionenuitwisselingschromatografie, voor het scheiden van de enzymen. Deze techniek scheidt ze van elkaar in de volgorde van de extractie Ι, ΙΙ en ΙΙΙ door de concentratie van ammoniumsulfaat te verhogen. De enzymen werden vernoemd naar de volgorde van extractie RNAP I, RNAP II, RNAP III.
Voor de transcriptie zijn een grote verscheidenheid aan transcriptiefactoren nodig voor het binden van de RNA-polymerase II aan de bovenstroomse promotors. Deze factoren vormen samen met de RNA-polymerase II het basale transcriptiecomplex.
Subeenheden
|
Saccharomyces cerevisiae Pol II subeenheden |
Al of niet in de andere RNA-polymerasen voorkomend |
Menselijk gen | Aantal proteïnogene aminozuren | Menselijke Pol II subeenheden |
|---|---|---|---|---|
| RPB1 | II | POLR2A | 1970 | subunit A |
| RPB2 | II | POLR2B | 1174 | subunit B |
| RPB3 | II, IV, V | POLR2C | 275 | subunit C |
| RPB4 | II | POLR2D | 142 | subunit D |
| RPB5 (RPABC1, ABC27) | I, II, III | POLR2E | 210 | hRPB5 (subunit E) |
| RPB6 (RPABC2, ABC23) | I, II, III, IV, V | POLR2F | 127 | hRPB6 (subunit F) |
| RPB7 | II | POLR2G | 172 | subunit G |
| RPB8 (RPABC3, ABC14,5) | I, II, III, IV, V | POLR2H | 149 | hRPB8 (subunit H) |
| RPB9 | II, IV, V | POLR2I | 125 | subunit I |
| RPB10 (RPB11-a) | II, IV, V | POLR2J | 117 | subunit J |
| RPB11 (ABC10β, RPABC4) | I, II, III, IV, V | POLR2K | 58 | hRPB10 (subunit K) |
| RPB12 (ABC10α, RPABC5) | I, II, III, IV, V | POLR2L | 67 | hRPB12 (subunit L) |
- DNA-directed RNA polymerase II subeenheid RPB1 – een enzym dat bij mensen het gen POLR2A afleest en bij gist RPO21. RPB1 is de grootste subeenheid van RNA-polymerase II. Het bestaat uit een carboxylterminaal eiwitmotief (CTD) met tot 52 heptapeptide repeats (Tyr-Ser-Pro-Thr-Ser-Pro-Ser), die onmisbaar zijn voor de polymerasewerking. Het CTD is ontdekt in het laboratorium van C.J. Ingles van de University of Toronto en door J.L. Corden van de Johns Hopkins University. In combinatie met verscheidene andere polymerase subeenheden vormt het RPB1 het DNA-bindingseiwitmotief, de groef waarin de DNA-template wordt getranscribeerd in RNA. Het heeft een sterke wisselwerking met RPB8.
- RPB2 (POLR2B) – de op een na grootste subeenheid, die in combinatie met ten minste twee andere polymerase subeenheden een structuur in de polymerase vormt, dat zorgt voor het contact van de actieve plaats van het enzym met de DNA-template en het nieuw gevormde RNA.
- RPB3 (POLR2C) – de op twee na grootste subeenheid. Het bestaat uit een heterodimeer met de polymerase subeenheid RPB10 en vormt een kernhalffabricaat. RPB3 heeft een sterke wisselwerking met RPB1-5, 7, 10-12.
- RPB4 – wordt afgelezen van het POLR2D-gen en speelt mogelijk een rol bij de bescherming tegen stress.
- RPB5 – wordt afgelezen van het POLR2E-gen. Twee moleculen van deze subeenheid komen voor in alle drie de RNA-polymerasen. RPB5 heeft een sterke wisselwerking met RPB1, RPB3 en RPB6.
- RPB6 (POLR2F) – vormt een structuur met ten minste twee andere subeenheden dat de transcriberende polymerase op de DNA-template stabiliseert.
- RPB7 – wordt afgelezen van het POLR2G-gen en speelt mogelijk een rol in de regulering van de polymerasewerking. RPB7 heeft een sterke wisselwerking met RPB1 en RPB5.
- RPB8 (POLR2H) – heeft een wisselwerking met de subeenheden RPB1-3, 5 en 7.
- RPB9 – De groef waarin de DNA-template wordt getranscribeerd in RNA is samengesteld uit RPB9 (POLR2I) en RPB1.
- RPB10 – wordt afgelezen van het POLR2L-gen. Het heeft een wisselwerking met de subeenheden RPB1-3 en 5 en een sterke wisselwerking met RPB3.
- RPB11 – de RPB11 subeenheid bestaat bij mensen uit de drie subeenheden: POLR2J (RPB11-a), POLR2J2 (RPB11-b) en POLR2J3 (RPB11-c).
- RPB12 – heeft ook een wisselwerking met RPB3 (POLR2K).
Vorming
RPB3 is betrokken bij de vorming van de RNA-polymerase II. Een subcomplex van RPB2 en RPB3 ontstaat spoedig na de vorming van de subeenheden. Dit complex reageert vervolgens met RPB1. RPB3, RPB5 en RPB7 vormen met elkaar homodimeren. Samen kunnen RPB3 en RPB5 reageren met alle andere RPB-subeenheden, uitgezonderd RPB9. Alleen RPB1 bindt sterk aan RPB5. De RPB1-subeenheid reageert ook met RPB7, RPB10 en zwakker maar meest efficiënt met RPB8. Pas als RPB1 aan het complex koppelt kunnen andere subeenheden zoals RPB5 en RPB7 ook aankoppelen, waarbij RPB5 bindt aan RPB6 en RPB8 en vervolgens RPB3 RPB10, RPB 11 en RPB12 aankoppelt. RPB4 en RPB9 koppelen pas aan als het grootste deel van het complex is gevormd. RPB4 vormt een complex met RPB7.
Werking bij mRNA
De verlenging van het mRNA door de transcriptie van de gekoppelde DNA-enkelstreng wordt gekatalyseerd door het RNA-Polymerase II.
De RNA-synthese kan worden gestart en de RNA-streng groeit in 5' → 3'-richting. Deze fase wordt elongatie genoemd. Tijdens de elongatie moet telkens het dubbelstrengs-DNA uit elkaar worden gehouden vlak voor de RNA-polymerase. Dit wordt door een transcriptiefactor bewerkstelligd, namelijk TFIIH (transcriptiefactor II met helicase activiteit). Vlak na de elongatie wordt aan het 5'-einde van de RNA-synthetiserende streng een beschermingselement aangebracht, een zogenaamde 'kap' ('cap').
RNA-polymerase II kan gefosforyliseerd (RNA-polymerase IIO) en niet gefosforyliseerd (RNA-polymerase IIA) zijn. De overgang van de ene naar de andere vorm maakt verschillende functies van de transcriptie mogelijk. De fosforylering van CTD wordt gekatalyseerd door een van de zes basale transcriptiefactoren, TFIIH. TFIIH heeft twee functies: een voor het ontwinden van het DNA op de plaats waar de transcriptie begint en een voor het fosforyleren. RNA-polymerase IIA hecht zich aan het pre-initiatiecomplex. RNA-polymerase IIO zorgt voor de verlenging van de RNA-streng. De verlenging begint met de fosforylering van serine op de positie5- (ser5), via TFIIH. het gefosforyliseerde ser5 zorgt ervoor dat enzymen het 5'-eind van het nieuw gevormde RNA kunnen cappen en de 3'-vormingsfactoren aan de poly(A)plaatsen kunnen vastmaken. As het tweede serine is gefosforyliseerd, ser2, wordt de verlenging geactiveerd. Defosforylisering zorgt voor het beëindigen van de verlenging. Als het eiwitmotief geheel is gedefosforyliseerd katalyseert RNA-polymerase II hetzelfde proces maar nu op een andere initiatieplaats.
In dit voorbeeld een ribulose-molecuul, dat al deel uitmaakt van het mRNA en de uracil-nucleotide UTP (uridinetrifosfaat):
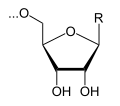 +
+ 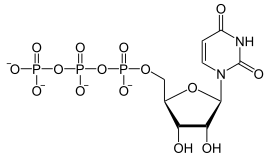 →
→  + PPi
+ PPi
De fosfaatgroep van het UTP wordt met de hydroxylgroep van het ribulose van het mRNA verestert. Het RNA-molecuul wordt verlengd onder afsplitsing van difosfaat (PPi).
Holo-enzym
RNA-polymerase II holo-enzym is een vorm van het eukaryotische RNA-polymerase II dat nodig is voor de promotoren van proteïne-coderende genen. Het bestaat uit RNA-polymerase II, een deel van de basale transcriptiefactoren en regulerende proteïnen, die SRB-proteïnen (Suppressor van RNA-polymerase B = onderdrukker van RNA-polymerase B) genoemd worden. (RNA-polymerase B = RNA-polymerase II).
Een deel van de vorming van het holo-enzym wordt het pre-initiatiecomplex genoemd, omdat de vorming plaatsvindt op de core-promotor van het gen voordat de transcriptie begint. De functie van dit complex is het correct positioneren van het RNA-polymerase II op de startpunten van de te transcriberen genen, denaturatie van het DNA en positionering van het DNA op de actieve plaats van het RNA-polymerase II.
Het carboxylterminaal eiwitmotief van RNA-polymerase
Het carboxylterminaaleiwitmotief (CTD = Carboxy Terminal Domain) van RPB1 bestaat uit tot 52 repeats met de sequentie Tyr-Ser-Pro-Thr-Ser-Pro-Ser. Het eiwitmotief loopt vanaf de kern van RNA-polymerase II tot aan het uitgangskanaal. Het CTD-eiwitmotief komt niet voor bij RNA-polymerase I en RNA-polymerase III. Het RNA-polymerase-CTD werd voor het eerst ontdekt in het laboratorium van C.J. Ingles van de University of Toronto en ook in het laboratorium van J Corden van de Johns Hopkins University tijdens het sequencen van het DNA, dat de RPB1-subeenheid van RNA-polymerase van gist en muis respectievelijk codeert. Andere eiwitten binden vaak aan het C-terminaaleiwitmotief van RNA-polymerase voor het activeren van de polymerase. Het carboxylterminaaleiwitmotief is betrokken bij het starten van de transcriptie, de capping van het RNA-transcript en het vastmaken aan de spliceosoom voor RNA-splicing.
Bepaling type RNA-polymerase
Alfa-amanitine wordt door zijn werkingsmechanisme ook veelal gebruikt als een stuk gereedschap in wetenschappelijke studies in moleculaire biologie en biologisch onderzoek. Het kan gebruikt worden om te bepalen welke vormen van RNA-polymerase aanwezig zijn. Men test dan de gevoeligheid van de RNA-polymerase in aanwezigheid van alfa-amanitine. RNA-polymerase I, RNA-polymerase IV en RNA-polymerase V zijn ongevoelig, RNA-polymerase II zeer gevoelig en RNA-polymerase III is enigszins gevoelig voor alfa-amanitine.
Externe links
| Bronnen, noten en/of referenties |